 WWU-Münster
WS 1998/99 WWU-Münster
WS 1998/99
Institut für Georaphie
Seminar: VEGETATIONSGEOGRAPHIE
Dozent: H. Schreiber
Standortfaktoren
biotische Faktoren
Protokoll vom 4.11.1998
Verfasser: Birgit Mikullo
und Daniel Bleyenberg
1. Einleitung
Tab.1
Gesamt-
wirkung |
Der Standort
als räumliche
Gegebenheit im Gelände ("Naturraum")
|
Faktoren-
komplex |
Klima
|
Relief
|
Boden
|
Biotische
Faktoren
|
| Einzel
Faktoren |
Strahlung
Niederschlags-
höhe
Schneedecke
Nebel
Temperaturen
Windverhältnisse
u.v.a. |
Hangneigung
Hangrichtung
Kleinrelief
Talgestaltung
Massenerhebung
u.a. |
Bodenart
Wassergehalt
Humusart und
-menge
Struktur
Kalkgehalt, pH
Nährstoffgehalt
Spurenelemente-
gehalt
Saltzgehalt
Grundgestein
Grundwasser-
stand
u.v.a. |
Boden-
organismen
pflanzliche
Konkurrenten
Wildtiere
Haustiere
Schädlinge
Maßnahmen des
Menschen
u.a. |
Wenn man bei Zeiten einmal
einen genaueren Blick auf die Parkanlagen, Klein- und Vorgärten wirft,
kann einem in den Sinn kommen, daß das Leben einer Pflanze recht
beschaulich sein muß. Ständig genug Licht und Wasser, für
Nährstoffe wird auch gesorgt, und gegen die lebensfeindlichen Temperaturen
im Winter bekommt man eine schützende Decke aus Tannenzweigen. Doch
in der Wildnis, abseits von Rasensprengern und Düngemitteln, ist das
Überleben nicht annähernd so unbeschwert... (Bem.: Kein Zitat,
sondern Vermutungen eines mitfühlenden Protokollanten!)
Aber bevor wir nun endgültig
in den Bereich der Komik abgleiten, kehren wir lieber wieder auf den rationalen
Boden der Vegetationsgeographie zurück. Der Lebenszyklus einer einzelnen
Pflanze und auch von großräumigen Gemeinschaften wird von zahlreichen
Faktoren beeinflußt (siehe Tab.1), die zusammengefaßt den sogenannten
Standort
ausmachen. Da diese Faktoren in einem engen Zusammenhang zueinander stehen
und sich somit gegenseitig beeinflussen und verändern, wächst
die Anzahl der verschiedenen Standorte praktisch gesehen bis ins Unendliche
- von dem einen Extrem der Wüste (minimale Feuchtigkeit) bis zum anderen
des Regenwaldes (maximale Feuchtigkeit). Ähnliche Beispiele lassen
sich auch für Wärme, Lichteinstrahlung und Bodenverhältnisse
anführen.
Aus so vielen Standort-Varianten
resultiert eine unglaubliche Artenvielfalt. Bis heute sind längst
nicht alle auf der Erde vorkommenden Arten erfaßt - vor allem im
tropischen Regenwald schlummert noch so manche Karriere für den einen
oder anderen Botaniker. Im Laufe der Evolution hat jede Pflanze ihr Äußeres
und ihre Funktionsweisen auf bestimmte Verhältnisse abgestimmt. Diese
Anpassung ist um so spezieller, je schwieriger die Bewältigung der
Umweltbedingungen ist.
Doch selbst wenn in Hinblick
auf das Klima, das Relief und den Boden (abiotische Faktoren) alles
geregelt ist, ist an ein unbeschwertes Wachstum noch lange nicht zu denken.
Denn zu jedem Standort gibt
es natürlich nicht nur eine Pflanze, sondern unzählige Arten,
die mit einander um die für sie idealen Bedingungen konkurrieren.
Es beginnt ein "Rennen" um Licht und Wasser, daß für gewöhnlich
der mit dem schnellsten Wachstum gewinnt. Die Verlierer müssen sich
mit dem Zufriedengeben, was für sie übrigbleibt, und stimmen
ihr Erscheinungsbild dann wiederum auf die neuen Verhältnisse ab.
Doch die Gewinner sind auch
noch nicht alle Sorgen los, denn Wildtiere und Parasiten leben meist auf
Kosten derer, die am meisten Energie zu vergeben haben.
All diese Faktoren, die
nicht aus "planetaren" Gegebenheiten resultieren (wie Klimaverhältnisse
und Bodeneigenschaften), fallen unter die Rubrik Biotische Faktoren
(siehe Tab.1). Anders als zu früheren Zeiten der Vegetationsgeographie
wird dieser Komplex mit zunehmender Intensität betrachtet, denn selbst
wenn alle andern Bedingungen stimmen, entscheidet zuletzt der biotische
Faktor, ob eine Art erfolgreich Wurzeln schlagen kann. Mit eben diesem
Thema beschäftigt sich dieses Protokoll. Der Einfluß des Menschen
auf den Naturraum wird hier allerdings noch nicht behandelt.

2. Energiehaushalt
Unter dem Aspekt des Parasitismus
ist in der Einleitung schon kurz der Begriff "Energie" gefallen. Damit
ist jene Energie gemeint, die ein Lebewesen (egal ob Tier oder Pflanze)
für seine Existenz benötigt: Zum einen die Bildung von physischer
Masse (bei Pflanzen Phytomasse), und zum anderen der Aufwand zur
reinen Lebenserhaltung. Beim Modell des Energiehaushalts ist die erstere
Form von entscheidender Bedeutung, da sie von einem Lebewesen an andere
weitergegeben werden kann.
Der Ursprung sämtlicher
Energie ist die Sonne. Mit Hilfe ihres Lichts kann eine Pflanze durch Photosynthese
aus Kohlendioxid und Wasser Glucose herstellen, die wiederum unerläßlich
für die Bildung von Phytomasse ist.
Diese Energieumwandlung
ist nur den Pflanzen möglich, weswegen diese Gruppe im Kreislauf den
Platz der
Produzenten einnimmt. Alle andern Lebewesen (in erster
Linie Tiere) beziehen ihre Kräfte von ihnen - entweder direkt durch
den Verzehr von Pflanzen, oder indirekt durch den Verzehr von Pflanzenfressern.
Die Beteiligten dieser Abfolge nennt man Konsumenten-Erster-Ordung,
Konsumenten-Zweiter-Ordung, u.s.w. Diese vermeintliche Einteilung ist
jedoch nicht unbedingt zwingend. So macht ein Fleischfresser hoher Ordnung
keinen großen Unterschied in der Wahl seiner Beute zwischen erster
oder zweiter Ordnung. Ebenso lassen sich die Allesfresser nicht eindeutig
in diese Systematik eingliedern.
Abb.1: Nährstoffkreislauf
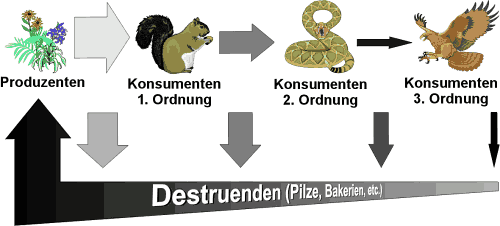
Weswegen trotzdem eine solche
Einteilung in verschiedene Trophieebenen (trophos gr. = Ernährung)
vorgenommen wird, liegt nun an der Energie, die im Kreislauf verloren geht
- eben jene Energie, die ein Tier für die Erhaltung seiner Körperfunktionen
benötigt, wie Blutkreislauf, Stoffwechsel oder Immunsystem. Die Folge
dieses Verlustes - er entspricht in etwa dem Faktor 10 - führt dazu,
daß die Anzahl der Exemplare bei ansteigender Trophiestufe
entsprechend abnimmt, da der Räuber ja nicht weniger Energie als seine
Beute fürs Überleben benötigt, sondern durch das Jagen eher
noch mehr. Es kann also eine gewisse Anzahl von Tieren erster Ordnung nur
eine viel geringere Zahl Tieren zweiter Ordnung ernähren. Jede Schwankung
in einer Population zieht gleichzeitig auch eine Veränderung in allen
anderen Ordnungen nach sich - positiv wie negativ.
Kreislauf bedeutet nun aber,
daß Energie nicht nur verbraucht wird, sondern dem Anfang (sprich:
den Produzenten) zurückgeführt wird. Für diesen Prozeß
sind die sogenannten
Zersetzer verantwortlich, oder auch Destruenten
genannt. Dabei handelt es sich um Bodentiere, Insekten, Pilze, Bakterien
und anderen Organismen, die Kot, Kadaver und abgestorbene Pflanzenteile
in Stoffe zersetzen, die Pflanzen wiederum aufnehmen und für ihr Wachstum
verwerten können. Da diese Zersetzung aber nicht 100%ig abläuft,
geht auch hier wieder Energie in Form von Dauerhumus verloren.
Der gesamte Verlust im Nährstoffkreislauf
durch Zersetzung und Lebenserhaltung gleicht die Sonne schließlich
wieder aus.
3. Bi-Systeme
Tab. 2:
|
Bi-Systeme
|
|
Bezeichnung
|
Wirkung:
A->B B->A |
Beispiele
|
|
Konkurrenz
|
| intraspezifische
Konkurrenz |
-/0
|
-
|
zwischen
Individuen gleicher Art |
| interspezifische
Konkurrenz |
-/0
|
-
|
zwischen
Individuen zweier Arten |
|
Antibiosen
|
| "Räuber-Beute" |
+
|
-
|
Herbivorie |
| Parasitismus |
+
|
-
|
Misteln,
Cuscuta, Orobanchen,
Würgefeige, Epiphylle,
Krankheiten |
|
Parabiosen
|
| Kommensalismus |
+
|
0
|
Pflanzen
des Waldbodens, Epiphyten |
| Amensalismus |
-
|
0
|
Heide-Bodenversauerung,
Laubstreu-
Moosschicht |
| Neutralismus |
0
|
0
|
so
gut wie nie vorhanden |
| Metabiosen |
+
|
0
|
Bodenverbesserung
durch
Pioniervegetation |
|
Sysmbiosen
im weiteren Sinn
|
| Allianz |
+
|
+
|
lockere Verbindung
zum
gemeinsamen Nutzen |
Mutualismus
(zeitlich
begrenzt, fakulativ) |
+
|
+
|
allgemeine Bestäubung
oder
Samenverbreitung |
| Symbiose im |
fakultativ |
+
|
+
|
Flechten,
Knöllchenbakterien,
Mycorrhiza, spezielle Bestäubung |
| engeren Sinn |
obligat |
+
|
+
|
Samenverbreitung,
Termiten-
Pizkulturen |
Unter diese Bezeichnung fallen
die Beziehungen zweier Lebewesen zueinander und den daraus resultierenden
Auswirkungen auf die weitere Entwicklung des einzelnen Individuums. Dabei
ist es egal, ob es sich um Tiere oder Pflanzen handelt.
Die Konkurrenz
ist schon in der Einleitung ein wenig vorgestellt worden. Der Wettbewerb
um den perfekten Standort ist natürlich energieaufwendig, weswegen
in dessen Verlauf keiner der beiden Beteiligten einen Gewinn verzeichnen
kann.
Es gibt aber auch Beziehungen,
bei denen ein Individuum auf Kosten eines anderen lebt, was nicht selten
in der Vernichtung desselben endet. So etwas fällt in den Bereich
der
Antibiosen.
Das Gegenteil davon sind
die Parabiosen, bei denen ein Individuum sich durch das andere
einen Vorteil verschafft, ohne es zu schädigen.
Und dann gibt es natürlich
auch die Symbiosen als Gegensatz zu der oben genannten Konkurrenz,
wo sich zwei Individuen gegenseitig unterstützen.
3.1. Konkurrenz
Wie bereits erwähnt
wurde, sind Konkurrenzverhältnisse für alle Beteiligten recht
ernergieaufwendig, was schließlich darin gipfelt, daß einer
von beiden nicht mehr genügend Kräfte für eine Vermehrung
mobilisieren kann. Ist es einmal so weit, bleibt dem Verlierer dieses Wettkampfes
nur noch der Rückzug zu einem anderen (etwas schlechteren) Standort,
oder aber er unterbricht seine Entwicklung und wartet an Ort und Stelle
auf seine Chance. Kommt die jedoch nicht, so wird auch dieses Individuum
irgendwann absterben.
Ein Unentschieden - bedeutet:
beide Pflanzen(-Arten) besitzen ein Gleichgewicht in ihren Energiekapazitäten,
was zu einer gegenseitigen Duldung führt - kommt in der Natur nur
recht selten vor, da der ausschlaggebende Umstand, der zwischen Sieg oder
Niederlage entschiedet, nur minimal sein muß. In der Regel nutzt
jede Art und Individuum jeden noch so kleinen Vorteil gegenüber anderen
sofort aus.
Bei der Konkurrenz wird
zwischen zwei Arten unterschieden:
Die intraspezifische
Konkurrenz bezieht sich auf den Wettbewerb unter Individuen gleicher
Art, während sich die interspezifische Konkurrenz
mit dem Verhältnis verschiedener Arten befaßt.
3.1.1.
intraspezifische Konkurrenz
Der innerartliche Wettbewerb
ist von den beiden Konkurrenzverhalten noch am einfachsten zu betrachten,
da man sich nicht mit den Eigenschaften mehrerer Arten auseinandersetzen
muß. Es wird mehr oder weniger davon ausgegangen, daß die zu
beobachtende Art sich bereits gegen andere Lebensformen durchgesetzt hat.
Die intraspezifische Konkurrenz ist demnach kein Vorgang, um andere Pflanzen
zu verdrängen, sondern um in der eigenen Art den kräftigsten
Individuum ein Überleben und deren Vermehrung zu sichern. "Es wird
also durch diesen Wettbewerb die Erhaltung der Art nicht gefährdet,
sondern gesichert." (Walter 1960: 16)
Wie so ein Konkurrenzkampf
in der Realität aussieht, läßt sich gut am folgenden Beispiel
verdeutlichen.

An diesem Beispiel (Fichte)
ist zu erkennen, daß bei weitem keine gleichmäßig bewachsene
Fläche herauskommen muß. Gerade an einem Berghang sorgen Wind-
und Frostschäden sowie Erdrutsche für einen immer wieder neu
aufflammenden Wettbewerb. Überall dort, wo genügend Freifläche
besteht, beginnen sofort Jungpflanzen zu wachsen. Und wenn sich ein Individuum
durchgesetzt hat, müssen die anderen ein Ausweichen versuchen, wodurch
diese Exemplare ihr Wachstum nicht nur nach oben, sondern auch zur Seite
ausrichten können. Aus demselben Grund ist auch die Ausbreitung des
Geästes eines Baumes und dessen Stammstärke in einer Gruppe selten
gleichmäßig. Die an den Seiten stehenden Pflanzen wollen das
hohe Lichtangebot natürlich nutzen, während den in der Mitte,
auf eine Weise auch geschützt stehenden Konkurrenten nur die Flucht
nach oben bleibt - sprich: Ast und Stammstärke müssen für
einen intensiven Wuchs in die Höhe herhalten.
Alleinstehende Individuen
sind allen Widrigkeiten (Wind und Schnee) am stärksten ausgesetzt
und müssen daher auch eine möglichst feste und kompakte Form
entwickeln.
Daß die Verlierer
der intraspezifischen Konkurrenz nicht immer gleich absterben müssen,
kann man am Beispiel der Buche (Fagus) sehen. So kann der Jungwuchs
über viele Jahre hinweg auf optimale Bedingungen warten. Eine kleine
Buche mit sehr dünnem Stamm kann daher nicht selten 150 und mehr Jahresringe
aufweisen - das genaue Alter läßt sich ohne Probleme durch die
Abnahme eines Bohrkerns bestimmen. Stirbt ein in der nähe stehendes
ausgewachsenes Exemplar ab, nimmt die Masse und Größe des Nachwuchses
rasant zu, bis sich am Ende wiederum ein Individuum durchgesetzt hat.
Wenn wir einmal bei dem Beispiel
der Buche bleiben, und uns den Wald ansehen, fällt einem auf, daß
außer ihr kaum noch andere Arten zu finden sind. Unterholz und Bodenbewuchs
ist nicht existent. Die Bedingungen für die Beobachtung einer interspezifischen
Konkurrenz, also der Wettbewerb zwischen verschiedenen Arten, sind daher
logischerweise nicht allzu optimal. Auf einem für sie idealen Standort
ist die Fagus sylvatica unschlagbar. Ihre Baumkronen sind derart
mächtig, daß für bodennahe Arten schlichtweg nicht genügend
Licht vorhanden ist.
Jenen Fall, wenn eine Pflanze
ideale physische (Licht, Wärme, Wasser, Boden,...) Bedingungen für
ihr Wachstum vorfindet, nennt man physiologisches Optimum.
Nun ist es interessant zu
beobachten, was passiert, wenn man Arten in ein Konkurrenzverhältnis
bringt, die - auf den ersten Blick - dasselbe physiologische Optimum besitzen.
In diesem Zusammenhang führten Walter, Ellenberg und weitere Kollegen
den sogenannten Hohenheimer Grundwasserversuch durch (siehe
Abb.3). Hierbei wird ein Becken mit konstantem Grundwasserspiegel mit Erde
aufgefüllt, deren Oberfläche jedoch eine gleichmäßige
Steigung erfährt. (Im ursprünglichen Versuch wurde die Füllung
zur Hälfte in Sand und Lehm geteilt, was jedoch auf das hier im Protokoll
beschriebene Ergebnis ohne großen Belang ist.) Das führt dazu,
daß die Feuchtigkeit der oberen Bodenschichten abnimmt - in der Graphik
von Links nach Rechts.
Der nun erläuterte
Versuch bezieht sich speziell auf das Zusammenleben von drei verschiedenen
Arten Wiesengras:
Alopecurus, Arrhenaterum, und Bromus.

[Walter, H. (1960)]
Wird jede Art für sich
alleine in Reinkultur gesät, so zeigt sich schnell, daß
sich ihr physiologisches Optimum im Bereich der mittleren Feuchtigkeit
befindet.
Als nächstes werden
dann alle drei Arten mehr oder weniger gleichmäßig in die Versuchsfläche
eingesät. Im Laufe der Zeit zeichnet sich ab, daß Arrhenaterum
kräftig genug ist, um den Bereich seines Optimums erfolgreich gegen
die Konkurrenz zu verteidigen. In den Extrembereichen verliert dieses Gras
jedoch seine Dominanz, da die beiden anderen Arten einen größeren
Spielraum in Puncto Feuchtigkeitsverträglichkeit besitzen. Also weichen
Alopecurus
und Bromus auf die von ihnen noch- tolerierbaren Randbereiche aus.
Ersteres flüchtet sich in die hohe Feuchtigkeit, während sich
das andere Gras in der Trockenheit durchsetzen kann. Allerdings ist ein
voll ausgeprägtes Wachstum in den Ausweichzonen nicht immer möglich,
da die Umweltbedingungen dort nur die zweite Wahl sind und nicht mehr hergeben.
Durch den Konkurrenzdruck
können die unterdrückten Arten also ihr physiologisches Optimum
nicht erreichen.
Da Konkurrenz als biotischer
Faktor die Qualität des Standortes mitbestimmt, wird die neue Situation
als
ökologisches Optimum bezeichnet. Die Durch-setzungsfähigkeit
einer Art hängt umgekehrt aber auch immer von den anderen Standortfaktoren
ab, wodurch man bei weitem nicht von einem konstanten Wettbewerb zweier
Arten untereinander ausgehen kann.
Wie das Ergebnis einer solchen
interspezifischen Konkurrenz aussieht, kann man im folgenden Beispiel eines
Ökogramms
(Abb.4) erkennen - hier für einen Mitteleuropa.
Im mittelfeuchten und schwach
sauren bis alkalischen Bereich ist die Rotbuche dominant. Andere Baumarten
haben es dort sehr schwer. Sie können erst dort eine Waldfläche
bilden, wo ihr Konkurrent durch die schlechteren Bodenverhältnisse
seine Stärke soweit verliert, daß sich andere Bäume behaupten
können. Doch auch dann findet sich immer eine Art, die in einem bestimmten
Bereich gegenüber ihren Konkurrenten den für sie optimalen Standort
vorfindet. [Ellenberg (1978)]
Abb.4: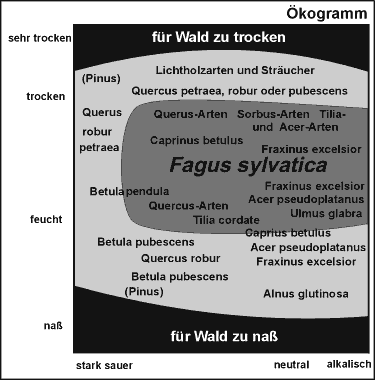
Nun gibt es aber auch Pflanzen,
die sich im Wettbewerb mit anderen Arten nicht nur auf schnelles Wachstum
und große Toleranzbreite verlassen, sondern sich auch (oder nur)
durch andere, mitunter recht aggressive Methoden Vorteile verschaffen.
So entwickeln die Blätter des Walnußbaums bei der Verrottung
toxische Stoffe, die die Nutzung des Bodens in ihrer Umgebung für
mögliche Konkurrenten unmöglich machen.
Mit fortschreitender Technik
hat die interspezifische Konkurrenz teilweise ganz neue Dimensionen erreicht.
Durch den regen kontinentalen Schiffsverkehr wurden Arten in Bereiche eingeschleppt,
die sie bislang durch natürliche Grenzen (Alpen oder Wüsten)
nie erreicht hätten. Zuerst in den Häfen vorkommend, können
sich einige Arten schließlich auch im großen Konkurrenzkampf
durchsetzten, was die Umgebung natürlich einigermaßen verändert,
indem heimische Arten einfach verdrängt werden.
Welche Folgen diese Vorgänge
haben können, zeigt das Beispiel der Spartina.
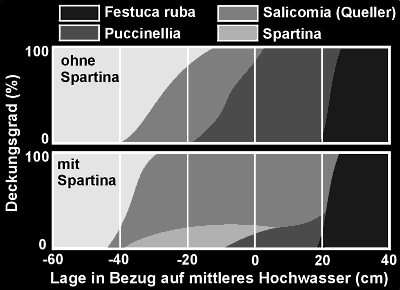
Abb. 5: Zonierung der dominierenden
Arten bei der Schlickablagerung
an der schleswig.holsteinischen
Nordseeküste (nach D.König, 1948)
An der Nordseeküste
Schleswig-Holsteins begann dieses Schlickgras allmählich das Andelgras
(Puccinellia) aus seinem ursprünglichen Lebensraum zu verdrängen,
was zunächst niemanden weiter störte, da die neue Pflanze scheinbar
ebenfalls recht gut die Schlickablagerungen befestigte. Der Haken daran
war nur, daß sie äußerst schlecht mit den Gezeiten zurechtkam,
was zu einer Auswaschung des Bodens führte, wodurch nun das Angelgras
keinen Halt mehr fand. Dem Vorgang der Landgewinnung wurde somit eine hilfreiche
Pflanze genommen.
3.2. Antibiosen
3.2.1. "Räuber-Beute"
Die Wirkungsweise zweier
Arten (bzw. eines Bi-Systems) kann nicht nur konkurrierend sein, sondern
u.a. antibiotisch. Als Beispiel wurde diesbezüglich aus dem Bereich
der Herbivoren (» Pflanzenfresser) das Eichhörnchen angeführt.
Es frißt die Samen der Eiche. Weil es sie aber auch verschleppt und
verteilt, um sich Nahrung für den Winter zu sichern, das Eichhörnchen
diese aber in seiner Geschäftigkeit oft vergißt, ist die Grenze
zu einer Symbiose mit der Eiche fließend, d.h. der Nachteil den die
Eiche durch die Antibiose hat, also durch den Fraß der Eichensamen
ist relativ gering i.V. zum Vorteil der Symbiose, durch die die Samenverteilung
durch das Eichhörnchen gewährleistet ist. Dieses Beispiel macht
deutlich, das es bei der Herbivorie, einem "Räuber- Beute" Verhältnis,
häufig nicht zu einer 100% Vernichtung kommt.
3.2.2. Parasitismus
So, wie das "Räuber-Beute"
Verhältnis den Antibiosen untergeordnet ist, läßt sich
auch der Parasitismus unterordnen, weil man unter Antibiosen ebenso folgendes
versteht: "i.w.S. die Beziehung v. Partnern verschiedener Artzugehörigkeit
zum Vorteil des einen u. Nachteil des anderen; die A. i. e. S. ist keine
bloße Nährstoffkonkurrenz, sondern wird durch besondere, v.
den Antibioten ausgeschiedene Stoffe (z. B. Antibiotika) verursacht." (Herder-Lexikon
der Biologie 1994, Bd. 1: 203).
Parasiten, auch "Schmarotzer"
genannt, lassen sich, in bezug auf die Nährstoffgewinnung, in Halb-
und Vollparasiten unterteilen. Als Halbparasit ist u. a. die
Mistel zu bezeichnen; sie besitzt noch grüne Organe, mit denen sie
einen Teil ihres Nährstoffbedarfs decken kann, den anderen Teil deckt
sie durch die Mitnutzung des Nährstoffaufnahmesystems ihres Wirts,
in den sie eingedrungen ist und an den sie sich festgeheftet hat. Die Halbschmarotzer
erstellen demnach einen Teil der benötigten Energie selbständig,
durch Photosynthese. Auch Orchideen sind Halbparasiten, sie "zapfen" ihren
jeweiligen Wirt an den Wurzeln an. Es gibt aber auch einige chlorophyllose
Orchideen (wie Corallorhiza), die gehören dann zu den Vollparasiten
(Holoparasiten).
Vollparasiten sind
demnach: "Pflanzen ohne Chlorophyll, die in ihrer Ernährung ganz auf
den Wirt angewiesen sind. Sie haben meist reduzierte Sprossen, Blätter
und Wurzeln, bilden dafür Saugorgane (Haustorien) aus, die bis zu
den Leitgefäßen des Wirts vordringen. Ihre Samen keimen oft
nur im Kontakt mit Wirtswurzeln" (Dierschke 1994: 46). Auf die Ficus
leprieurie (eine Feigenart: Würgefeige) weisen auch Michael G.
Barbour u. a. in ihrem Buch "Terrestrial Plant Ecology" (1987) als einen
Vollparasiten hin, der nach und nach seine Wirtspflanze bis zur vollständigen
Überdeckung in Beschlag nimmt.
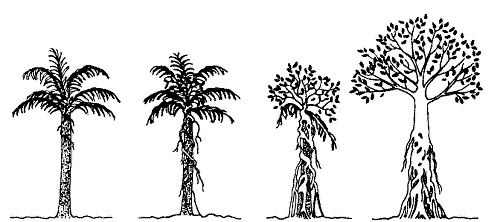 Abb.6: Parasitismus
der Würgefeige
Abb.6: Parasitismus
der Würgefeige
Mit der Abbildung 6 ( "Figure
7-4" ;Barbour, M. G. et al; 1987; S.137) wurden vier Phasen dieses Prozesses
deutlich gemacht: Bild (a) zeigt, daß die Samen auf den Baum gelangt
sind und auskeimen; auf Bild (b) sieht man die Keimlinge herunterwachsen;
Bild (c) zeigt, daß der Kontakt zum Boden aufgenommen worden ist,
was ein Dickenwachstum der Pflanze zu Folge hatte und einen Ausbau der
Krone bewirken konnte; Bild (d) zeigt das Erwürgen des Wirts, der
Baum verrottet vollständig, Hohlräume sind dadurch entstanden,
aber die Feige ist mittlerweile so dick, daß sie alleine stehen kann.
Die Würgefeige kann somit als echter Parasit bezeichnet werden.
Loranthacee (Mistelgewächse)
werden ebenso als Halbparasiten gezählt; sie setzen sich auf Bäume
und verbinden in raffinierter Weise den Parasitismus mit dem Epiphytismus.
Krankheiten, die von Viren und Bakterien verursacht werden, stellen ebenfalls
Antibiosen dar.
Allgemein wird die Produktion
und Abgabe der für andere Organismen schädlichen Stoffe als Antibiose
bezeichnet. So bilden manche Pflanzen Giftstoffe in ihren Blättern,
um im Falle eines "Angeknabbert werdens", einerseits ihren Artgenossen
auf diese chemische Weise eine bevorstehende "Gefahr" zu signalisieren
(Hilfeleistung), andererseits um diesen Giftstoff selber als Abwehrstoff
zu nutzen.
In Anlehnung an Tabelle 2,
an das Bi-System, gehen wir als nächstes auf die Parabiosenein.
Als Formen der Parabiose werden hier Kommensalismus, Amensalismus,
Neutralismus und Metabiosen genannt.
3.3.1. Kommensalismus
Unter Kommensalismus versteht
man laut Herder eine "Form des Zusammenlebens artverschiedener Tiere, bei
der die eine, meist kleiner Art (Kommensale) v. der Nahrung der anderen
Art (Wirt) profitiert, ohne den Partner zu schädigen od. ihm zu nützen"
(Herder-Lexikon der Biologie 1994, Bd. 5: 78). Als Beispiel wurden Epiphyten
(genauer Flechten) genannt, z.B. Flechten, die sich auf Fichten niederlassen
oder auf anderen Bäumen; sie sitzen meist in Zweigtrichtern, um dort
das angesammelte Regenwasser zu trinken; sie zersetzen hier Organisches
und entnehmen die Nährstoffe (Epiphytismus). Auch die Buche wurde
als Beispiel angeführt: Sie nimmt durch ihre große Krone, den
Bodenpflanzen das meiste Licht; für andere Pflanzen am Waldboden,
die wenig Licht brauchen, aber viel Platz, ist dies ein willkommender Standort.
3.3.2.
Amensalismus
Mit Amensalismus bezeichnet
man "eine Art des Zusammenlebens zweier Arten a und b, bei der a einen
negativen Einfluß auf b, aber b keinen Einfluß auf a ausübt"
(Herder - Lexikon der Biologie 1994, Bd. 1:143).
Bei der Überlegung,
wo es Amensalismus denn gäbe, kam man auf die Bodenversauerung durch
das Heidekraut zu sprechen, was als ein Beispiel in Frage kam. Das Heidekraut
muß H+-Ionen an den Boden abgeben, damit es Nährstoffe
aufnehmen kann, das führt zu einer Bodenversauerung und stellt eine
indirekte Konkurrenz für andere Arten dar, die dort bald nicht mehr
wachsen können. Moose stehen im Buchenwald unter einem negativen Einfluß.
Weil der Boden durch schwer zersetzbares Laubstreu der Buche bedeckt wird,
muß die Moosdecke aufgrund des entstandenen Lichtmangels teilweise
weichen - sie wird schütterer.
3.3.3. Neutralismus
In bezug auf den Neutralismus
kann angemerkt werden, daß dieser so gut wie nicht existiert, da
Pflanzen auf einem Standort eigentlich immer auf irgendeine Art und Weise
Einfluß aufeinander ausüben - wenn auch nicht unbedingt radikal.
Dennoch läßt sich für ein sich gegeseitig ignorierendes
Verhältnis zwischen verschieden Arten kaum ein Beispiel finden (was
im Seminar auch nicht gefunden wurde).
3.3.4. Metabiosen
Zu Metabiosen halten
wir fest, daß der Nutzeffekt meist durch eine Zeitverschiebung gekennzeichnet
ist, beispielsweise können an Küstengebieten, aufgrund des Antragens
von Sand durch das Meer, Pionierpflanzen den Boden vorbereiten, damit zu
einem späteren Zeitpunkt sich andere Pflanzenarten dort verbreiten
können, eine Bodenverbesserung ist damit einhergehend (veränderte
Standorte).
3.4. Symbiosen im weiteren
Sinn
Unter dem Punkt "Symbiosen
im weiteren Sinn" wurden in erwähnter Tab.2 als weitere Formen
Allianz, Mutualismus und Symbiosen im engeren Sinne (fakultativ / obligat)
genannt. Auf die Form der Allianz, laut Abb. 3, eine lockere Verbindung
zum gemeinsamen Nutzen, wurde in der Sitzung nicht weiter eingegangen.
Anders verhielt sich das beim Mutualismus (auch Eusymbiose genannt).
Auch diese Form ist wie jede andere der Symbiose "eine Plus-Plus-Beziehung
zum Vorteil beider Partner" (Glavac 1996: 61), die zusätzlich ein
zeitlich begrenztes Zusammensein beinhaltet. Sie stellt die "engste Form
wechselseitiger, lebensnotwendiger Abhängigkeit mit gegenseitig günstigen
Wirkungen" dar (Dierschke 1994: 46). Dieses Gleichgewicht zwischen Angriff
und Abwehr bewirkt eine erhöhte Konkurrenzkraft gegenüber anderen
Organismen. Als Beispiele von Mutualismus galten hier die allgemeinen Vorgänge
der Bestäubung und Samenverbreitung wie z.B.
(A) auf der Salbeipflanze
wird die Hummel, die auf Nahrungssuche ist, nach innen geleitet, dabei
drückt sie auf ein Widerlager, was dazu führt, daß der
Stengel Pollen auf dem Hinterteil der Hummel ablegt; die Pollen mit Hilfe
der Hummel so zu anderen Pflanzen gebracht
(B) der Schlangenwurzel
entwickelt übelriechende Duftstoffe und lockt so Fliegen an, die die
Bestäubung übernehmen; die Fliegen fallen in die Röhre und
bekommen Pollen aufgedrückt- dann ziehen sie weiter zum nächsten.
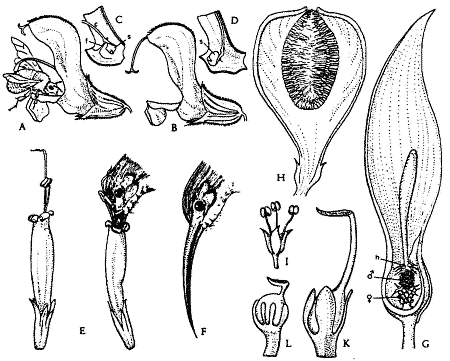
Abb. 7 :Tierblütigkeit
bei verschiedenen Angiospermen. A-D Hummel als Blütenbesucher an Salvia
pratensis (violettblau) (etwas vergr.). E-F Der Honigvogel Arachnothera
longirostris als Bestäuber bei Sanchezia nobilis (Acanthaceae,
Blüten gelb, Brakteen purpurn) etwa ¾). G Aufgeschnittener
Blütenstand (Gleitfallenblume) von Arum maculatum mit hellgrüner
Spatha und unscheinbaren m,w und Hindernisblüten (h) im weiblichen
Entwicklungszustand (2/3 X). H Blütenstand von
Ficus carica im Längsschnitt (etw. vergr.) mit männl. (I) und
langgriffeligen weibl. (K) fertilen Blüten sowie kurzgriffeligen weibl.
Gallenblüten (L) (vergr.). [Strasburger (1978), S. 759]
Außerdem gehen Gefäßpflanzen
vielfältige mutualistische Beziehungen mit Rhizobium und anderen
Mikroorganismen ein (Glavac 1996: 61). Unter Rhizobium versteht
man Knöllchenbakterien, die bestimmte Pflanzen (Hauptwirte), wie z.B.
Erbse, Saatwinkel, Weißklee, Bohne, Luzerne, Steinklee oder Hornklee,
über deren Wurzelhaare infizieren und dort Stickstoff (N2)
fixieren.
3.4.1. Symbiosen im engeren
Sinn
Zu Symbiosen im engeren
Sinn (fakultativ / obligat) eignen sich besonders Flechten (Lichenen)
als Beispiele. Sie sind durch eine hochentwickelte Symbiose zwischen Pilzen
und Algen charakterisiert. Bei ihnen gibt es nur artspezifische Symbiosen,
die i. d. R. von Außen nicht erkennbar ist.
Der Pilz erhält von
der Alge Kohlenhydrate, die Alge ist in der Umhüllung durch das Pilzgeflecht
geschützt, z. B. vor raschem Wasserverlust. Die Symbiose ermöglicht
den Partnern das Vorkommen an Standorten, die sie alleine vielfach nicht
besiedeln könnten (Herder-Lexikon der Biologie 1994, Bd.: 340). Im
Groben: Der Pilz stellt also die Masse dar, die Feuchtigkeit halten kann
und die Alge ist für den Gewinnungsvorgang bei der Photosynthese verantwortlich.
Ein weiteres Beispiel für
eine Symbiose ist die Mycorrhiza. Darunter versteht man eine Symbiose
zwischen einem Pilz und einer höheren Pflanze (Nultsch - Allgemeine
Botanik, 1991: 349). Zur Veranschaulichung dieser Symbiose dient die Abbildung
8 "Mycorrhiza" (eine RE-Aufnahme aus dem Internet), welche eine Ektomycorrhiza
zeigt. Hier umspinnt ein dichtes Pilzmycel mantelartig die keulig angeschwollenen
Wurzelenden (von Dryas octopetala?) (Nultsch - Allgemeine Botanik, 1991:
349, 350). Der Vorteil der Pflanze besteht in der Fähigkeit des Mycorrhizapilze,
Mineralstoffe für sie verfügbar zu machen, dafür erhält
der Pilz vor allem Kohlenhydrate. Beide ziehen also einen Vorteil aus dieser
Beziehung.
Abb. 8: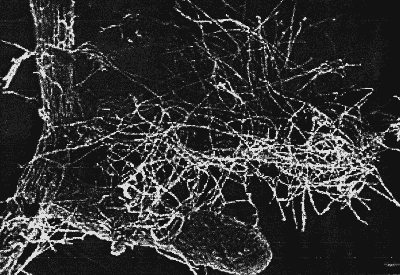
Das das Prinzip der Symbiose beschänkt
sich natürlich nicht nur auf den Bereich der Flora, sondern ist auch
zwischen Pflanzen und Tieren üblich.
4. Abschluß
Alle hier im Protokoll beschriebenen
Bi-Systeme sind nicht unter dem Gesichtspunkt des Menschen betrachtet worden.
Doch gerade die Auswirkungen unserer Zivilisation hat unverkennbare Veränderungen
in Sachen Standort hervorgerufen: Nitrate, Saurer-Regen, Grundwasser-verbrauch
und andere Eingriffe in den globalen Naturraum haben die Voraussetzungen
für ein ungehindertes Wachstum vieler Pflanzen teilweise drastisch
verschlechert. Konkurrenzkämpfe, die längst beendet schienen,
keimen erneut auf. Das Gesicht ganzer Vegetationszonen wandelt sich - Beispiel:
Regenwald.
Für die Zufunft der
Georgraphen und Botaniker verspricht dies zwar äußerst gute
Beschäftigungsaussichten, auf die sie aber wohl gut und gerne verzichten
könnten.
5. Literatur-Verzeichnis
[1] Barbour, M. G.,
Burk, J. H. & Pitts, W. D. (1980): Terrestrial plant ecology. 1.Aufl.
Menlo park
[2] Dierschke, H.
(1994): Pflanzensoziologie. Stuttgart: Eugen Ulmer GmbH & Co.
[3] Ellenberg, H. (1978):
Vegetation Mitteleuropas mit den Alpen in ökologischer Sicht, 2.Aufl.,
Stuttgart
[4] Glavac, V. (1996):
Vegetationsökologie. Jena: Gustav Fischer Verlag
[5] Herder (1994):
Lexikon der Biologie. Bd 1-9. Heidelberg: Spektrum Akademischer Verlag
GmbH. Sauermost, Rolf (Hrsg.)
[6] Klink H.-J. (1996):
Vegetationsgeographie, 2.Aufl., Braunschweig (=Das Georgraphische
Seminar)
[7] König, D.
(1948): Spartina townsendii an der Westküste Schleswig-Holsteins,
Planta (Berl.)
[8] Nultsch, W. (1991):
Allgemeine Botanik. 9., neubearb. Aufl. Stuttgart: Georg Thieme Verlag
[9] Schreiber, H.J. (1998):
Die Waldgrenznahen Buchenwälder und Grasländer des Falakron und
Pangäon in Nordostgriechenland. Synataxonomie, Struktur und Dynamik,
Münster (=Arb. Inst. Landschaftsökol. Bd.4)
[10] Strasburger, E.
(1978): Lehrbuch der Botanik. 31.Aufl., Stuttgart, S. 759
[11] Walter, H. (1960):
Einführung in die Phytologie Bd. 3: Grundlagen der Pflanzenverbreitung,
1.Teil: Standortslehre, 2.Aufl., Stuttgart |